Primary Research Interests
Selected features of the Neurospora genome/epigenome (LGII)
H3K9me3 directs DNA methylation
Our genetic and biochemical studies on the control of DNA methylation revealed ties between DNA methylation and chromatin modifications. We discovered that the DIM-2 (defective in methylation) DNA methyltransferase is directed by heterochromatin protein 1 (HP1), which in turn recognizes trimethyl-lysine 9 on histone H3 (H3K9me3), placed by the DIM-5 histone H3 methyltransferase. DIM-5 is sensitive to modifications of histones (notably, methylation of Lysine 4 and phosphorylation of Serine 10 of histone H3) and is found in a complex (DCDC) with four other proteins that are all essential for DNA methylation: DIM-7, DIM-8 (DDB1), DIM-9 and CUL4 (see figure below). We are anxious to determine the mechanism that directs DCDC to regions destined to be constitutive heterochromatin, such as the centromere regions and other regions characterized by degenerate DNA sequences left by the genome defense mechanism RIP (Repeat-induced point mutation). RIP is a homology-based premeiotic process that litters duplicated sequences with C:G to T:A mutations.
RIP searches the genome for repeated sequences (e.g. transposons) in the sexual phase of the life cycle between fertilization and nuclear fusion and causes polarized transition mutations and DNA methylation.
We have found that a variety of factors modulate DNA methylation. For example, mutants in dmm-1 (DNA methylation modulator-1) show aberrant methylation of DNA and histone H3K9, with both frequently spreading into genes adjacent to inactivated transposable elements, leading to poor growth. Mutants defective in both dmm-1 and dim-2 display normal H3K9me3 patterns, implying that the spread of H3K9me3 involves DNA methylation but, in general, HP1 and DIM-2 are dispensable for H3K9me3. We have identified several silencing complexes that involve HP1, most notably the histone deacetylase complex HCHC, which is illustrated in the figure below.
We are also investigating additional factors controlling DNA methylation, including DIM-1, DIM-3, SET-3, CDP-2, among others. Further, we have identified DNA methylation-independent factors that are important for silencing of genes associated with constitutive heterochromatin and are anxious to explore their mode of operation.
Heterochromatin machinery of Neurospora.
Histone H3 Lysine 27 methylation (H3K27me)
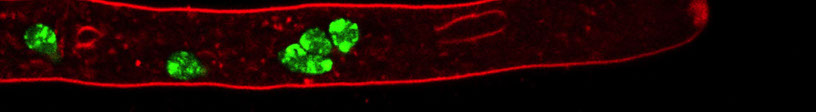
Work in flies, mammals and plants implicated H3K27me in long-term repression of genes, including X-inactivation and cancer in mammals. The mechanisms responsible for localization of H3K27me are largely unknown, in part because of the complexity of epigenetic regulatory networks. As part of our program using N. crassa to elucidate the control and function of H3K27me, we are exploring interactions between constitutive and facultative heterochromatin. H3K27me normally covers ~7% of the Neurospora genome, encompassing hundreds of transcriptionally silent genes. The Polycomb repressive complex 2 (PRC2) subunits SET-7, EED and SU(Z)12 are each required for H3K27me, while the fourth PRC2 subunit, NPF is only needed for H3K27me near the ends of chromosomes. We found that elimination of heterochromatin protein 1 (HP1) or any member of the DCDC H3K9 methylation complex (which generates the "mark" that HP1 binds to in constitutive heterochromatin) cause massive redistribution of H3K27me to regions that are normally marked by H3K9me3; in contrast, elimination of DNA methylation by deletion of the DNA methyltransferase gene has no effect on H3K27me (e.g. see figure).
Relocalization of H3K27me2/3 occurs in H3K9me3 mutant but not in DNA methylation mutant
We are using HiC to interrogate interactions of sequences across the genome in wildtype and mutant strains of Neurospora to investigate nuclear organization and its control (sample result shown in Circos plot). Surprisingly, even in the absence of H3K9me3 or HP1, constitutive heterochromatin regions show strong interactions and these interactions are facilitated by Importin α (DIM-3).
Results of HiC on wildtype Neurospora shows strongest interchromosomal (inside circle) and intrachromosomal (outside circle) interactions occur between constitutive heterochromatin sequences marked by H3K9me3 (green) or between facultative heterochromatin marked by H3K27me2/3 (purple)